
Формирование половых продуктов птиц. Зародышевая линия организма представляет собой популяцию клеток, которые в своем развитии дают начало формированию половых клеток, сперматозоидов и яйцеклеток. Жизненный цикл линии зародышевых клеток начинается от оплодотворения. Также как и у других позвоночных, у птиц закладка зародышевой линии происходит во внегонадной области на ранних стадиях развития. У птиц, в частности у кур, на X стадии развития, что соответствует свежеснесенному яйцу, в бластодерме имеется немногочисленная (до 30-40 шт.) популяция клеток, обладающая полипотентными свойствами. Эти клетки локализованы в центральной области бластодермы [6].
style="display:inline-block;width:336px;height:280px"
data-ad-client="ca-pub-4037835599918832"
data-ad-slot="7553000704">
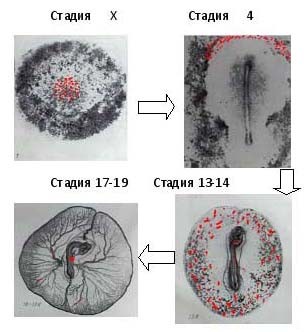

В случае самки, ПЗК, попавшие в левый яичник, дифференцируются для формирования оогоний после 8 дней инкубации и начинают активно делиться в процессе дифференциации в первичные ооциты. К 16 дням инкубации первичные ооциты входят в профазу мейоза. Возобновление развития первичного ооцита происходит при достижении половозрелости. Первичный ооцит выталкивает первое полярное тельце в результате первого мейотического деления примерно за 1 час до овуляции и превращается во вторичный ооцит. В это время определяется, какую из хромосом Z или W содержит вторичный ооцит, фактически происходит генетическое определение пола эмбриона. После овуляции происходит оплодотворение, и вторичный ооцит выталкивает второе полярное тельце и становится зрелой яйцеклеткой. 4-20 сперматозоидов проникают в яйцеклетку и каждый из них формирует мужской пронуклеус. Один из них сливается с женским пронуклеусом и становится оплодотворенным яйцом (4 часа после овуляции). Гонады позвоночных состоят из двух различных компонентов. Строма формируется из целомического эпителия и тканей мезонефроса, в то время как гаметы формируются из первичных зародышевых клеток (ПЗК). Во что разовьются ПЗК — в спермии или ооциты не зависит от их генотипа (т.е. от ZZ или ZW), а определяется типом стромы, в котором они развиваются. У нормальных самок птиц развивается левый яичник, а правый остается рудиментом. Удаление левого яичника приводит к пролиферации правого яичника, но уже как семенника, продуцирующего сперму из ПЗК самок.На стадии Х ПЗК обнаруживаются с помощью иммуногистохимического анализа с использованием моноклональных антител SSEA-1 и EMA-1. Из вентральной поверхности зоны пеллюцида на стадии Х1-Х1V они постепенно переходят в вентральную область зоны гипобласта. К 18 часам инкубации (стадия 4 по Гамбургеру и Гамильтону [10]) ПЗК перемещаются вперед, в район зародышевого полумесяца (фиксируются как йодная кислота-Шифф реакция позитивные). Оттуда они попадают в кровоток, некоторое время находятся в кровотоке и проникают в гонады (см. рис.1). ПЗК, проникшие в гонады, в случае самца, начинают активно делиться после 13 дней инкубации и дифференцироваться в сперматогонии.
Кагами и др. [11] в 1995 году сообщили, что ПЗК генетических самцов птиц могут дифференцироваться в функционирующие яйца, а ПЗК генетических самок — в функционально нормальные сперматозоиды. Это было показано путем анализа судьбы бластодермальных клеток самок и самцов, инъецированных в реципиентные эмбрионы противоположного пола.
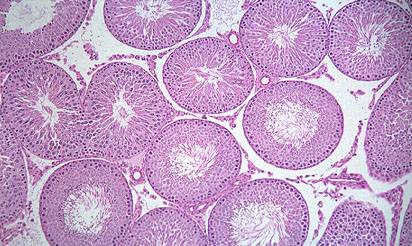
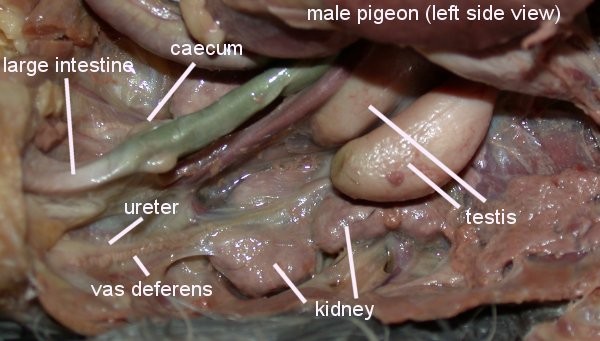
Созревание спермы и половая система самца. Созревание спермы происходит в семенных канальцах семенников (рис.2) Этот процесс лучше происходит при низких температурах, поэтому сперматогенез преимущественно имеет место в ночное время суток, когда температура тела ниже.
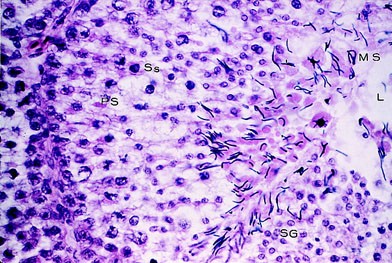


Сперма хранится в удаленном конце семенного канатика (vas deference, seminal glomus), и это создает возвышение, называемое клоакальным бугорком. Самцы птиц имеют парные абдоминальные семенники, занимающие краниовентральное положение к первой доле почек (рис. 3, 4). Размеры семенников быстро увеличиваются в ходе племенного сезона. Vas deferens проходит каудально к клоаке, где имеет общий с мочеточником выход в часть клоаки, называемый уродеум (Urodeum). Терминальный конец vas deferens набухший вследствие хранения спермы: семенной клубок (или семенной пузырек).
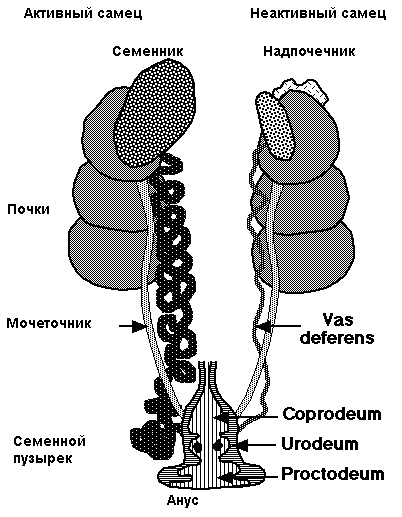

Как и у млекопитающих, у которых развита мошонка, содержащая семенной клубок (внешняя терморегуляция), формирование спермы у птиц — процесс зависящий от температуры, и ее созреванию способствует ночное падение температуры.
Кроме того, самцы птиц имеют сравнительно маленький запас внегонадной спермы и она эякулируется вскоре после формирования в семенниках.
Конкуренция спермы и размеры семенников.
Сравнительные анализы показывают, что различные экологические и поведенческие факторы оказывают влияние на значительную вариабельность по количеству создания половых пар среди птиц. При анализе 1010 видов птиц, Pitcher et al. [13] проверили несколько экологических и поведенческих факторов в связи с размерами семенников; индекс конкуренции спермы и уровень создания половых пар среди птиц.

В одномерных и многомерных анализах, размер семенников у колониальных видов был значительно больше, чем у тех, которые создают одиночные пары, что говорит о том, что высокая плотность размножения связана с большей конкуренций спермы. После проверки филогенетических влияний и других экологических переменных, размер семенников был также больше в таксонах, которые не участвуют в кормлении своих потомков. При анализе данных по различиям, как по групповым видам, так и по показателю филогенетической независимости, у моногамных таксонов семенники меньше, чем у таксонов с многочисленными социальными связями, и размер семенников увеличивается с увеличением размера кладки, что свидетельствует о том, что истощение спермы могло играть роль в эволюции размеров семенников. Данные результаты свидетельствуют о том, что традиционные экологические и поведенческие переменные, такие как система социальных связей, плотность размножения и отцовская забота самца могут объяснить значительную долю отклонений по показателю конкуренции спермы у птиц.
Формирование яйца и половая система самки. У большинства птиц функционирует только один яичник и один яйцевод. На ранних стадиях эмбрионального развития, каждая самка имеет по два яичника; только левый развивается в функционирующий орган. У некоторых птиц, таких как ястребы, обычно развиваются и правый яичник и яйцевод. Зрелый яичник выглядит как гроздь и может содержать до 4000 маленьких яиц, которые могут развиваться в зрелые фолликулы (рис.5) .
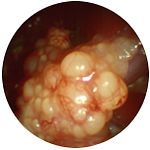
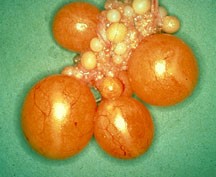




Рисунок 5. Яичник птиц (слева — незрелые яйца, в середине — развитые фолликулы, справа — яичник, яйцевод и яйцо со скорлупой).
В сезон размножения яичник значительно увеличивается. Активные яичники схожи на грозди винограда — развивающиеся фолликулы. Яйцевод открывается воронкой медиально к яичнику (рис. 6).
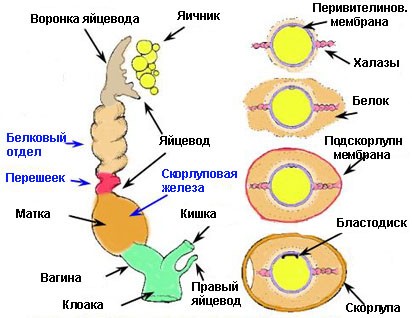

Овуляция приводит к освобождению яйца из зрелого фолликула на поверхности яичника. Яйцо, с большим количеством запасных питательных веществ в форме концентрических слоев желтка, захватывается воронкой и потоком ресничек направляется в белковый отдел яйцевода. Более трех часов яйцо окружается белком. Яйцо затем поступает в перешеек, где формируются подскорлупные мембраны. На это уходит около одного часа. Яйцо затем движется в матку или
скорлуповую железу, где добавляется известковая скорлупа и, у некоторых птиц, добавляется пигмент. Яйцо затем проходит во влагалище и клоаку для откладки.
Самки могут влиять на пол своих цыплят. У некоторых видов птиц, пол птенца определяется условиями окружающей среды, в частности, обеспеченностью кормовыми ресурсами. Rutstein et al. [14] регулировал прием пищи у самок и самцов зяблика и установил, что при хорошем кормлении самки больше склонны давать дочерей, в то время как при плохом кормлении — сыновей. Такой результат может исходить из того факта, что потомство самок необходимо кормить лучше, чем самцов для выживания и роста.
Авторы отмечают, что у многих животных соотношение полов приближается к 50:50 и очень тяжело изменить его. У млекопитающих, включая человека, пол ребенка определяется тем, какая половая хромосома в сперме. Но у птиц, яйцо самки, а не сперма самца определяет пол цыпленка. Таким образом, самка определяет, яйцо самки или самца овулируется. В некотором роде, самка зяблика, казалось бы, может повлиять на пол птенцов в зависимости от того, какой из них имеет больше шансов на выживание. «Исследования говорят нам, что они это делают, и мы понимаем почему. Основной вопрос заключается в том: как они это делают».
У многих животных, самки нуждаются в хорошем кормлении и хороших условиях, если они должны давать потомство, так как яйца стоят очень дорого для организма. Из больших яиц вылупляются большие птенцы, у которых больше шансов на выживание. Такое регулирование соотношения пола отмечается у некоторых насекомых, таких как пчелы, осы, но значительно менее понятно у птиц и млекопитающих.
Птицы являются хорошей моделью для изучения регулирования полового соотношения, так как пол птенцов можно определить сразу после отложения яйца при помощи молекулярных методов. Более того, все ресурсы, необходимые для развивающегося эмбриона, находятся в яйце при отложении. Таким образом, размер и содержимое яйца являются мерой запасных веществ, которых самка вложила в яйцо, что в свою очередь влияет на последующее выживание.
Определение пола у птиц. Молекулярные механизмы, лежащие в основе определения пола у птиц, до сих пор остаются загадочными. Этот процесс отличается от такового у млекопитающих. У птиц не найдены гены гомологичные таковым, какие отвечают за самцовый тип у млекопитающих. Эта ситуация, вероятно, является отражением того факта, что, несмотря на присутствие двух полов практически во всем животном царстве, задействованные гены практически не связаны у разных классов многоклеточных организмов. Эти различия вызывают трудности для сравнительного анализа кандидатных генов в исследованиях полового развития [5].
У птиц самки — гетерогаметный пол, с одной копией Z и W половых хромосом. Самцы гомогаметны (ZZ). Однако, не ясно, то ли присутствие W хромосомы, характерной для птиц, запускает развитие самки, или же доза Z хромосомы определяет самца. Еще одной интригующей возможностью является то, что и Z, и W хромосомы играют роль в определении пола птицы! Например, у сумчатых животных, Y действует как хромосома, определяющая семенники, в то время как, X хромосома определяет выбор между мешком и мошонкой. Возможно, у птиц тоже используется система, где две половые хромосомы разделяют различные аспекты половой дифференциации.
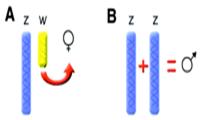

Температура инкубации и соотношение полов у птиц. Хотя у рептилий это и обычное явление, температура инкубации считается фактором, не оказывающим влияние на соотношение полов у птиц. Однако, Goth и Booth [8] обнаружили, что температура инкубации действительно оказывает влияние на соотношение полов у сорных кур (megapodes), какие отличаются от остальных птиц тем, что используют тепло из окружающей среды для инкубации яиц. У австралийских сорных кур (Alectura lathami), строящих кучи, при низких температурах вылупляются самцы, а при высоких — самки, в то время как при средней температуре в кучах в природных условиях соотношение равно 1:1. Цыплята при низких температурах весят меньше, что, вероятно, сказывается на выживаемости потомства. Megapodes имеют гетероморфные половые хромосомы, как и другие птицы, что исключает температуро — зависимое определение пола, какое описано у рептилий, как механизма, смещающего половое соотношение при высоких и низких температурах. Goth и Booth предполагают наличие температуро — чувствительной смертности эмбрионов, поскольку смертность была выше при низких и высоких температурах, и минимальной при средней температуре, при которой половое соотношение было 1:1.
Осеменение и оплодотворение. Самцы у некоторых видов птиц, включая большинство водоплавающих и страусов, имеют орган, приспособленный для введения спермы в половые пути самки. Большинство птиц не имеют фаллоса и у этих видов осеменение происходит через так называемый 'поцелуй клоаки'. В зависимости от таксона, сперма эякулируется в клоаку или вагину и, благодаря своей подвижности, достигает многочисленных канальцев для хранения, расположенных на границе между маткой и влагалищем. Вследствие селекции в ходе миграции через вагину, только 1-2% осемененной спермы входит в канальцы, остальные, вероятно, выбрасываются во время дефекации самки. Канальцы содержат только морфологически нормальную сперму, что указывает или на то, что только нормальная сперма способна пересечь вагину, или, что только нормальная сперма 'принимается' канальцами. Точно не известно, но небольшое количество спермы движется к воронке яйцевода (место оплодотворения), не входя в канальцы, хотя они, вероятно, оплодотворяют только одно яйцо [1].
Вся сперма в канальцах располагается головками, направленными в сторону дистального конца канальца что означает, что выход из них является пассивным. Сперма теряется из канальцев более или менее постоянно с определенной скоростью. Сперма входит в матку и пассивно передвигается к воронке. Она аккумулируется или медленно движется через воронку так, что обычно всегда имеется популяция сперматозоидов, готовая оплодотворить каждое яйцо при овуляции. После овуляции яйцо захватывается воронкой, приспособленной для этого, и множество сперматозоидов окружает поверхность яйца (рис. 9); их цель — зародышевый диск, который содержит женский пронуклеус. На этой стадии яйцо окружается внутренними перивителиновыми слоями (IPVL). Скопления спермы и дырки, проделанные в IPVL вокруг зародышевого диска свидетельствуют о том, что сперма, возможно, использует химические сигналы для нахождения зародышевого диска.


В отличие от большинства других таксонов, где только один спермий проникает в яйцо, у птиц полиспермия — обычное явление. Несколько спермиев проникают в район зародышевого диска, растворяя IPVL при помощи акросомной реакции спермия. Освобождение ферментов из акросомы спермия дает возможность ядру спермия проникнуть в яйцо.
Однако только один сперматозоид сливается с пронуклеусом самки, а остальные спермии смещаются к периферии зародышевого диска и дальше в развитии не участвуют. Оплодотворение включает проникновение спермия в яйцеклетку и слияние мужского и женского пронуклеусов (syngamy). Поскольку развитие эмбриона начинается почти сразу же, ко времени окружения яйца яйцевыми оболочками зародыш состоит из большого количества клеток.

King et al. [12] обнаружили, что сперматозоиды из двух различных осеменений (одно с окрашенной, другое с неокрашенной спермой) обычно изолируются в различные канальцы как у кур, так и у индеек. Канальцы, содержащие смешанные популяции сперматозоидов были найдены только у 4% цыплят и 12% индеек. Они заключили, что не срабатывает механизм преимущества последнего самца благодаря отсутствию наслоения сперматозоидов в канальцах.
Freedman et al. [7] проведя иммуногистохимическое окрашивание влагалищно-маточного соединения у индейки, обнаружил нейроны, иннервирующие поверхностный эпителий, выстилающий просвет влагалищно-маточного соединения канальцев для хранения спермы. Более того, авторы обнаружили присутствие актина (протеин, характерный для гладких мышц) в области канальцев. Эта связь между нейронами и канальцами для хранения спермы говорит в пользу того, что канальцы иннервируются и что хранение и выделение спермы из них может контролироваться.
Птичье яйцо. Яйца птиц, также как и сами птицы сильно различаются по размерам. Самые крупные яйца откладывают страусы. Их размеры в 2000 раз больше, чем самые маленькие яйца колибри. Яйца страусов имеют 180 мм в длину и 140 мм в ширину и весят 1.2-1,6 кг. Яйца колибри- 13 мм в длину и 8 мм в ширину и весят только пол грамма. Исчезнувшая птица-слон из Мадагаскара откладывала яйца в 7 раз крупнее, чем страус! Внутри яйца жизнь эмбриона обеспечивают три внеэмбриональные мембраны: амнион, хорион, аллантоис (рис. 10).
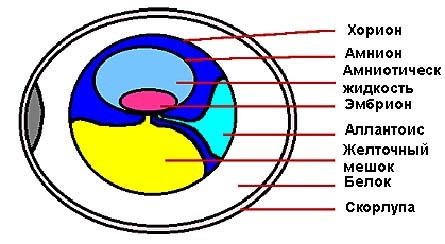

Амнион окружает только эмбрион. Внутренний слой клеток амниона секретирует жидкость, в которой плавает эмбрион. Амниотическая жидкость предохраняет эмбрион от высыхания и защищает его.
Хорион окружает все эмбриональные структуры и служит в качестве защитной мембраны.
Аллантоис — вырост задней кишки, становится шире по мере роста эмбриона, сливаясь с хорионом, формирует хориоаллантоисную мембрану, работает совместно с хорионом, обеспечивая дыхание и выделение, служит хранилищем для азотистых отходов (мочевая кислота).
Яйца состоят из 4-х первичных компонентов:
1. желток, богатый энергией источник питания, состоит из 21 — 36% липидов и 16 — 22% протеинов (остальное-вода). Желток висит в центре яйца на двойных протеиновых канатиках (халазах).
Состав яйца и фенотип цыплят. Вклад родителей в яйца и, следовательно, в потомство, может значительно влиять на фенотип, выживаемость и эволюционную приспособленность организма. Яйца птиц — прекрасная модельная система для изучения материнского энергетического вклада, переданного через вариации размеров яйца. Dzialowski1и Sotherland [3] использовали природный разброс по размерам яйца у эму (Dromaius novaehollandiae), от 400 г до >700 г, для проверки влияния материнского вклада в яйца на морфологию и физиологию новорожденных страусят.
Самки эму снабжают более крупные яйца большим количеством энергии, питательными веществами и водой в желтке и белке. Различия в материнском вкладе отражались в различиях размеров страусят, которые увеличивались с размерами яйца. Размер яиц также повлиял на физиологию растущих эмбрионов; так, уровень метаболических реакций эмбриона находился в позитивной связи с размерами яйца и эмбрионы, развивающиеся в крупных яйцах, потребляли больше желтка в ходе развития. Из больших яиц вылуплялись более тяжелые страусята (чистый вес без желтка и сухая масса) и структурно крупнее.
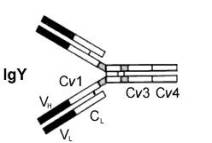
Желток содержит материнские антитела. Антитела откладываются в яйце во время формирования желтка в виде его иммуноглобулинов, сначала — IgY (также называемый IgG) (рис. 11).

У цыплят (Gallus domesticus), материнские IgY используются потомством в первые 14 суток жизни и к 5 суткам после лупки цыплята начинают синтезировать свои собственные IgY. В результате, после около двух недель жизни, циркулирующие IgY, у цыплят практически имеют эндогенное происхождение. Взрослых уровней титры антител достигают между 6 неделями и 6 месяцами жизни. Однако материнские антитела могут и дальше продолжать оказывать влияние на скорость роста и развития. В отсутствие материнских IgY у цыплят (вследствие хирургического удаления бурсы у матери в ходе ее эмбриогенеза), количество клеток в селезенке, которые помогают лимфоцитам (хелперные T клетки) уменьшается, атака антигенов (чужеродные протеины на патогенах) подавляется. Также подавлен иммунный ответ потомства, что может привести к снижению выживаемости потомства [9].
Почему желток желтый (или красный)? Желток яйца окрашивается каротиноидами. До недавнего времени не было объяснения, почему яйцекладущие животные так обильно насыщают яйца каротиноидами. Оказывается, что в развивающемся организме каротиноиды предохраняют уязвимые ткани от разрушений, вызываемых свободными радикалами. Хотя эмбриональные ткани и зависят от окисляемых, ненасыщенных жирных кислот в желтке, их избыток делает ткани восприимчивыми к перекисному окислению, вызываемому химически активными окислительными метаболитами и свободными радикалами, которые продуцируются как нормальные промежуточные продукты метаболизма. Защита против перекисного окисления липидов у цыплят осуществляется каротиноидами желтка и другими антиоксидантами, такими как витамин E. Антиоксиданты также защищают пассивно приобретенные антитела (IgY) от разрушения. Таким образом, материнский вклад в состав яйца, включая каротиноиды, имеет значительное влияние на выживаемость потомства. [2].
2. Белок состоит на 90% из воды и на 10% из протеина, выполняет роль поставщика воды для эмбриона, служит также для защиты эмбриона от резких перепадов температуры.
3. Подскорлупные мембраны. Две подскорлупные мембраны защищают эмбрион от проникновения бактерий и помогают предотвратить быстрое выпаривание влаги из яйца.


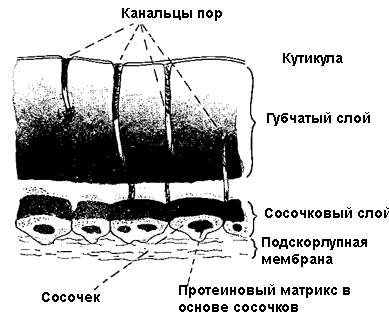
4. Скорлупа защищает эмбрион, содержит тысячи пор (рис. 13), которые обеспечивают газовый обмен. Скорлупа яиц обычно белая у птиц, гнездящихся в полостях (норах, дуплах) и цветная или узорчатая у птиц с открытыми гнездами. Цвет скорлупы приобретается благодаря пигментам, секретируемым клетками в стенке матки.
Слабые птицы используют стероиды для помощи потомству. Verboven et al. [16] сообщили, что самки слабых чаек оказывают гормональную поддержку для увеличения выживаемости своих птенцов. Verboven и ее коллеги экспериментально усилили материнский эффект при помощи дачи кормовых добавок во время формирования яйца и сравнили концентрации стероидов (включая тестостерон) в их яйцах с контролем. Андрогены яйца могут влиять на продуктивность потомства непосредственно через развитие цыпленка и/или опосредованно через изменение их конкурентоспособности по сравнению с собратьями. Против ожидания, самки с экспериментально усиленным рационом, откладывали яйца с низким уровнем андрогенoв. Это говорит о том, что менее здоровые самки передают больше стероидов, чем здоровые, чтобы усилить их производительность. Verboven заметила, что «Мы сначала думали, что чайки при хорошем питании отложат больше стероидов в своих яйцах. Но обнаружили, что здоровые птицы не передают в свои яйца излишнюю поддержку».

Она сравнивает ситуацию с борцами-атлетами, принимающими стероиды для увеличения силы и добавляет: «Слабые спортсмены, возможно, хотят использовать стероиды для нивелирования своей слабости, но если вы в хорошем состоянии, то вам и не нужно принимать их».
Самки птиц создают различные фенотипы путем различных гормональных вкладов в свои яйца. У птиц самки откладывают значительные количества андрогенов в свои яйца и экспериментальные данные показывают, что материнские андрогены оказывают влияние на раннее развитие цыпленка. Несмотря на хорошо известную организационную роль половых стероидов на мозг и поведение, работы в этой области ограничены в основном стадией цыпленка. Eising et al. [4] обнаружили, что, у чайки-черноспинки, материнские андрогены в яйце усиливают развитие брачного оперения и частоту агрессивных и половых проявлений почти год спустя после лупки. Этот продолжительный эффект может быть опосредован регуляцией рецепторов андрогенов на поздних этапах жизни. Или же ранняя гормональная обработка могла влиять на ось: гипоталамус — гипофиз — гонады, приведя к усилению продукции андрогенов во взрослой жизни. Эти долговременные эффекты андрогенов яйца также благотворны для дарвиновской адекватности. Удачное занятие территории и ее защита с помощью агрессивного поведения важны для успеха воспроизводства у колониальных птиц. Кроме того, эти проявления важны для выбора партнера. Ясно, у птиц материнский гормональный вклад в яйца может значительно влиять на индивидуальные различия признаков связанных с приспособлением. Поскольку эти гормоны подавляют раннюю иммунную функцию цыпленка и снижают продолжительность жизни, матери сталкиваются с компромиссом между производством потомков с низкой выживаемостью, но высоким репродуктивным потенциалом на год, или высокие шансы на долгожительство, но низкий ежегодный репродуктивный потенциал. Путем производства яиц, которые отличаются по уровню материнских гормонов, самки, вероятно, производят различные фенотипы, что, возможно является адаптивной стратегией в непредсказуемых условиях окружающей среды. Поскольку природная селекция действует при таких фенотипических вариациях, формируя демографию популяции, роль материнских андрогенов в этом селекционном процессе может быть значительно больше, чем считалось раньше.
Список литературы
1. Birkhead, T. R. and J. -P. Brillard. Reproductive isolation in birds: postcopulatory prezygotic barriers. // Trends in Ecology and Evolution. -2007. -V. 22.
2. Blount J. D., Houston D. C. and Muller A. P. Why egg yolk is yellow. // Trends in Ecology and Evolution. — 2000. -V. 15. -P. 47-49.
3. Dzialowskil, E. M. and P. R. Sotherland. Maternal effects of egg size on emu Dromaius novaehollandiae egg composition and hatchling phenotype. // J. Exp. Biol. — 2004. -V. 207 -P. 597-606.
4. Eising, C. M., W. Muller and T.G.G. Groothuis. Avian mothers create different phenotypes by hormone deposition in their eggs. // Proc. Royal Soc. London, Biol. Letters. -2005.
5. Ellegren, H. Hens, cocks and avian sex determination: a quest for genes on Z or W? // European Molecular Biology Organization Reports -2001. -V.2. -P. 192-196.
6. Eyal-Giladi H., Kochav S. From cleavage to primitive streak formation: A complementary normal table and a new look at the first stages of the development of the chick I. General morphology // Developmental Biology. — 1976. -V. 49 — P. 321-337.
7. Freedman, S. L., V. G. Akuffo, and M. R. Bakst. Evidence for the innervation of sperm storage tubules in the oviduct of the turkey (Meleagris gallopavo). // Reproduction. -2001. -V.121 -P. 809-814.
8. Goth, A. and D. T. Booth. Temperature-dependent sex ratio in a bird. // Biology Letters -2005. -V.1 -P. 31-33.
9. Grindstaff, J. L., E. D. Brodie III, and E. D. Ketterson. Immune function across generations: integrating mechanism and evolutionary process in maternal antibody transmission. // Proc. Royal Soc. Lond. -2003. V. 270 -P. 2309—2319.
10. Hamburger V., Hamilton H. L. A series of normal stages in the development of the chick embryo. // Morphology. -1951. -V.88, -P. 49-92.
11. Kagami H., Clark M.E., Verrinder Gibbins A.M. et. al. Sexual differentiation of chimeric chickens containing ZZ and ZW cells in the germline // Molecular Reproduction and Development. — 1995. -V. 42. — P. 379-387.
12. King L.M., J. P. Brillard, W.M. Garrett, et. al. Segregation of spermatozoa within sperm storage tubules of fowl and turkey hens. // Reproduction. -2002. -V.123 -P.79-86.
13. Pitcher T. E., P. O. Dunn and L. A. Whittingham. Sperm competition and the evolution of testes size in birds. // Journal of Evolutionary Biology -2005. -V.18 -P. 557-567.
14. Rutstein A.N., P. J. B. Slater, and J. A. Graves. Diet quality and resource allocation in the zebra finch. // Proc. R. Soc. Lond. B (Suppl.). -2004. -V. 20.
15. Samour Jaime H. The Reproductive Biology of the Budgerigar (Melopsittacus undulatus): Semen Preservation Techniques and Artificial Insemination Procedures. // Journal of Avian Medicine and Surgery. -2002. -V. 16 -P. 39-49.
16. Verboven N., P. Monaghan, D.M. Evans et.al. Maternal condition, yolk androgens and offspring performance: a supplemental feeding experiment in the Lesser Black-backed Gull (Larus fuscus). // Proceedings of the Royal Society: Biological Sciences. -2003. -V. 270 -P. 2223 — 2232.