Формирование половых продуктов птиц. Зародышевая линия организма представляет собой популяцию клеток, которые в своем развитии дают начало формированию половых клеток, сперматозоидов и яйцеклеток. Жизненный цикл линии зародышевых клеток начинается от оплодотворения. Также как и у других позвоночных, у птиц закладка зародышевой линии происходит во внегонадной области на ранних стадиях развития. У птиц, в частности у кур, на X стадии развития, что соответствует свежеснесенному яйцу, в бластодерме имеется немногочисленная (до 30-40 шт.) популяция клеток, обладающая полипотентными свойствами. Эти клетки локализованы в центральной области бластодермы [6].
style="display:inline-block;width:336px;height:280px"
data-ad-client="ca-pub-4037835599918832"
data-ad-slot="7553000704">
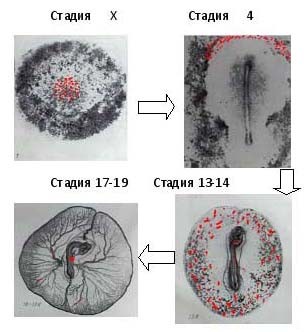
В случае самки, ПЗК, попавшие в левый яичник, дифференцируются для формирования оогоний после 8 дней инкубации и начинают активно делиться в процессе дифференциации в первичные ооциты. К 16 дням инкубации первичные ооциты входят в профазу мейоза. Возобновление развития первичного ооцита происходит при достижении половозрелости. Первичный ооцит выталкивает первое полярное тельце в результате первого мейотического деления примерно за 1 час до овуляции и превращается во вторичный ооцит. В это время определяется, какую из хромосом Z или W содержит вторичный ооцит, фактически происходит генетическое определение пола эмбриона. После овуляции происходит оплодотворение, и вторичный ооцит выталкивает второе полярное тельце и становится зрелой яйцеклеткой. 4-20 сперматозоидов проникают в яйцеклетку и каждый из них формирует мужской пронуклеус. Один из них сливается с женским пронуклеусом и становится оплодотворенным яйцом (4 часа после овуляции). Гонады позвоночных состоят из двух различных компонентов. Строма формируется из целомического эпителия и тканей мезонефроса, в то время как гаметы формируются из первичных зародышевых клеток (ПЗК). Во что разовьются ПЗК — в спермии или ооциты не зависит от их генотипа (т.е. от ZZ или ZW), а определяется типом стромы, в котором они развиваются. У нормальных самок птиц развивается левый яичник, а правый остается рудиментом. Удаление левого яичника приводит к пролиферации правого яичника, но уже как семенника, продуцирующего сперму из ПЗК самок.На стадии Х ПЗК обнаруживаются с помощью иммуногистохимического анализа с использованием моноклональных антител SSEA-1 и EMA-1. Из вентральной поверхности зоны пеллюцида на стадии Х1-Х1V они постепенно переходят в вентральную область зоны гипобласта. К 18 часам инкубации (стадия 4 по Гамбургеру и Гамильтону [10]) ПЗК перемещаются вперед, в район зародышевого полумесяца (фиксируются как йодная кислота-Шифф реакция позитивные). Оттуда они попадают в кровоток, некоторое время находятся в кровотоке и проникают в гонады (см. рис.1). ПЗК, проникшие в гонады, в случае самца, начинают активно делиться после 13 дней инкубации и дифференцироваться в сперматогонии.
Кагами и др. [11] в 1995 году сообщили, что ПЗК генетических самцов птиц могут дифференцироваться в функционирующие яйца, а ПЗК генетических самок — в функционально нормальные сперматозоиды. Это было показано путем анализа судьбы бластодермальных клеток самок и самцов, инъецированных в реципиентные эмбрионы противоположного пола.